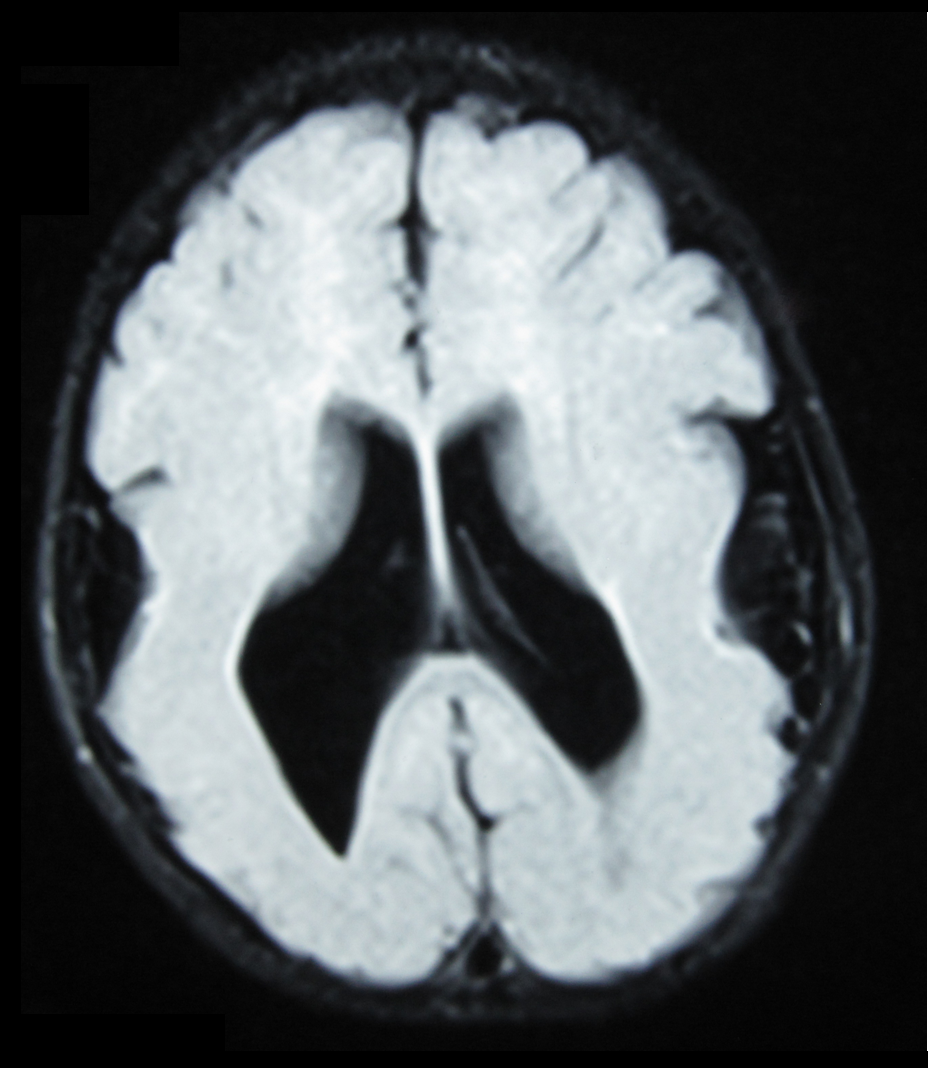
In the Dark of the Womb, a Thousands-Kilometer Journey in Micrometers
Imagine: a single neuroblast — small, colorless, nameless — is born in the lateral ventricle of a human embryo. It's not a mature neuron yet; it doesn't have an axon, hasn't formed synapses. But in its DNA, an ancient navigation map is stored: direction, distance, chemical signals, and a biological clock that leads its journey. Within 12-20 days, it must travel
1.5 to 3 millimeters — a distance equivalent to 500 times its body length — towards the cerebral cortex. There, it will line up, differentiate, and become one of the 86 billion neurons that form our soul.
But what if the map is blurry? If chemical signals — like reelin, LIS1, or DCX — fail to convey the message? Then the cell stops halfway. Or turns around. Or settles in the wrong layer. And it's not just one or two cells — but thousands, tens of thousands, even a large part of the cortical neuron population. That's where Neuronal Migration Disorder (NMD) is born: not because the brain doesn't grow, not because cells die, but because they arrive at the wrong destination, with precision, with full force — only in the wrong place, not where evolution intended.
When the Brain is Built 'Correctly' but in the Wrong Place
This is the core paradox of NMD — and why it's so hard to detect in routine ultrasounds or early MRIs. Babies with NMD often are born with a normal-sized head, non-suspicious muscle tone, and intact neonatal reflexes. No blood, no tumors, no trauma. Just: a thick layer of cortex where it should be thin; nodules of neurons clumped under the ventricle like lost islands; or a layer of cortex that's completely missing — replaced by an abnormal layered structure called
cobblestone lissencephaly.
Doctors don't see 'damage'. They see overly regular organization — but in a location that defies all principles of brain architecture. Like a skyscraper built perfectly… on the ocean floor. All the bricks are there, all the pillars stand, all the floors are complete — only it's not on solid ground.
Names that Hide the Horror: Lissencephaly, Polymicrogyria, Heterotopia
These terms are not just clinical labels — they are the epic geography of neurology.
Lissencephaly ('smooth brain') refers to a cortex without folds — no gyri, no sulci — because neurons failed to reach the brain surface.
Polymicrogyria, on the other hand, is the opposite: too many fine folds, too dense, because neurons stopped too early and clumped together excessively. And
subcortical heterotopia — the most poignant name — means 'a cluster of alien neurons under the cortex': like a colony of refugee nerve cells stranded in the subcortical region, waiting for a call that never comes.
Each name is a mirror to a migration tragedy that occurred between the 6th and 24th week of pregnancy — a phase that cannot be monitored directly, cannot be intervened, and cannot be repeated.
Behind the Silent Genes: Reelin, LIS1, and the Lost Language
We now know: NMD is not just bad luck. It's a mistake in the
molecular language. The reelin protein — released by Cajal-Retzius cells — acts like a 'signpost' for crossing neurons. Mutations in the
RELN gene cause neurons to stop in the inner layer, like a traveler who loses GPS in the forest. The
LIS1 gene, on the other hand, controls microtubules — the 'highway' within cells where neurons glide. When LIS1 is weak, neurons lose their drive, like a ship without wind.
What's most touching: children with DCX mutations — a gene that's only active in males — can have normal IQ, warm social skills, and a radiant smile… but never walk without support or speak more than ten words. Their brain is capable, but not connected — not because of a lack of intelligence, but because that intelligence is scattered in inaccessible places.
Not a Final Diagnosis — But a Starting Point to Understand Humanity
NMD teaches a quiet yet profound truth: that human identity is not just about
what is in the brain, but
where it is. Language is not just neuronal activity — it's the result of precise spatial coordination between Broca's area, Wernicke's area, and the connecting pathways that must traverse specific microscopic distances. Movement is not just motor impulse — it's the synchronization between the motor cortex, basal ganglia, and cerebellum, all of which depend on their relative positions.
The story of NMD is not a story of biological failure. It's a story of neuroplasticity's tenacity — how a 'lost' brain still tries to build new roads, form alternative synapses, and shift functions to more flexible areas. And in every smile, every attempt to reach out, every second of eye contact that persists against all odds — lies a miracle: life rewriting itself, letter by letter, neuron by neuron, even without the original map.
We no longer ask, 'What's wrong?'
We ask: 'Where does it differ — and what can we learn from it?'
Because sometimes, the answer to the most human question — 'Who am I?' — lies not in what's perfect, but in how something 'lost' still finds its way to speak.
This Baby's Brain Cells Moved in the Wrong Direction — and the Science World Now Understands Why. In the womb, a baby's nerve cells move like an ancient caravan — traversing microscopic distances that determine their lifelong fate. But what happens when their direction is wrong? It's not just a 'defect', but a condition where the brain is built *correctly* — just in the *wrong place*. This is not a neuroscientific fantasy: it's real, documented, and changes the way we look at human intelligence, movement, and language.. In the Dark of the Womb, a Thousands-Kilometer Journey in Micrometers
Imagine: a single neuroblast — small, colorless, nameless — is born in the lateral ventricle of a human embryo. It's not a mature neuron yet; it doesn't have an axon, hasn't formed synapses. But in its DNA, an ancient navigation map is stored: direction, distance, chemical signals, and a biological clock that leads its journey. Within 12-20 days, it must travel 1.5 to 3 millimeters — a distance equivalent to 500 times its body length — towards the cerebral cortex. There, it will line up, differentiate, and become one of the 86 billion neurons that form our soul.
But what if the map is blurry? If chemical signals — like reelin, LIS1, or DCX — fail to convey the message? Then the cell stops halfway. Or turns around. Or settles in the wrong layer. And it's not just one or two cells — but thousands, tens of thousands, even a large part of the cortical neuron population. That's where Neuronal Migration Disorder NMD is born: not because the brain doesn't grow, not because cells die, but because they arrive at the wrong destination , with precision, with full force — only in the wrong place, not where evolution intended.
When the Brain is Built 'Correctly' but in the Wrong Place
This is the core paradox of NMD — and why it's so hard to detect in routine ultrasounds or early MRIs. Babies with NMD often are born with a normal-sized head, non-suspicious muscle tone, and intact neonatal reflexes. No blood, no tumors, no trauma. Just: a thick layer of cortex where it should be thin; nodules of neurons clumped under the ventricle like lost islands; or a layer of cortex that's completely missing — replaced by an abnormal layered structure called cobblestone lissencephaly .
Doctors don't see 'damage'. They see overly regular organization — but in a location that defies all principles of brain architecture. Like a skyscraper built perfectly… on the ocean floor. All the bricks are there, all the pillars stand, all the floors are complete — only it's not on solid ground.
Names that Hide the Horror: Lissencephaly, Polymicrogyria, Heterotopia
These terms are not just clinical labels — they are the epic geography of neurology. Lissencephaly 'smooth brain' refers to a cortex without folds — no gyri, no sulci — because neurons failed to reach the brain surface. Polymicrogyria , on the other hand, is the opposite: too many fine folds, too dense, because neurons stopped too early and clumped together excessively. And subcortical heterotopia — the most poignant name — means 'a cluster of alien neurons under the cortex': like a colony of refugee nerve cells stranded in the subcortical region, waiting for a call that never comes.
Each name is a mirror to a migration tragedy that occurred between the 6th and 24th week of pregnancy — a phase that cannot be monitored directly, cannot be intervened, and cannot be repeated.
Behind the Silent Genes: Reelin, LIS1, and the Lost Language
We now know: NMD is not just bad luck. It's a mistake in the molecular language . The reelin protein — released by Cajal-Retzius cells — acts like a 'signpost' for crossing neurons. Mutations in the RELN gene cause neurons to stop in the inner layer, like a traveler who loses GPS in the forest. The LIS1 gene, on the other hand, controls microtubules — the 'highway' within cells where neurons glide. When LIS1 is weak, neurons lose their drive, like a ship without wind.
What's most touching: children with DCX mutations — a gene that's only active in males — can have normal IQ, warm social skills, and a radiant smile… but never walk without support or speak more than ten words. Their brain is capable , but not connected — not because of a lack of intelligence, but because that intelligence is scattered in inaccessible places.
Not a Final Diagnosis — But a Starting Point to Understand Humanity
NMD teaches a quiet yet profound truth: that human identity is not just about what is in the brain, but where it is. Language is not just neuronal activity — it's the result of precise spatial coordination between Broca's area, Wernicke's area, and the connecting pathways that must traverse specific microscopic distances. Movement is not just motor impulse — it's the synchronization between the motor cortex, basal ganglia, and cerebellum, all of which depend on their relative positions.
The story of NMD is not a story of biological failure. It's a story of neuroplasticity's tenacity — how a 'lost' brain still tries to build new roads, form alternative synapses, and shift functions to more flexible areas. And in every smile, every attempt to reach out, every second of eye contact that persists against all odds — lies a miracle: life rewriting itself, letter by letter, neuron by neuron, even without the original map.
We no longer ask, 'What's wrong?'
We ask: 'Where does it differ — and what can we learn from it?'
Because sometimes, the answer to the most human question — 'Who am I?' — lies not in what's perfect, but in how something 'lost' still finds its way to speak.